
Bordetella pertussis
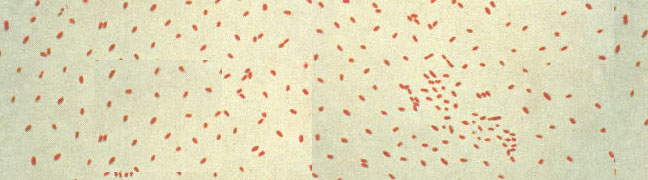
Whooping cough (pertussis) is caused by the bacterium Bordetella pertussis, B. pertussis is a very small Gram-negative aerobic coccobacillus that appears singly or in pairs. Its metabolism is respiratory, never fermentative, and taxonomically, Bordetella is placed among the "Gram-negative Aerobic Rods and Cocci" in Bergey's Manual. Bordetella is not assigned to any family. The bacteria are nutritionally fastidious and are usually cultivated on rich media supplemented with blood. They can be grown in synthetic medium, however, which contains buffer, salts, an amino acid energy source, and growth factors such as nicotinamide (for which there is a strict requirement). Even on blood agar the organism grows slowly and requires 3-6 days to form pinpoint colonies.
Bordetella pertussis colonizes the cilia of the mammalian respiratory epithelium (Figure 1). Generally, it is thought that B. pertussis does not invade the tissues, but some recent work has shown the bacterium in alveolar macrophages. The bacterium is a pathogen for humans and possibly for higher primates, and no other reservoir is known. Whooping cough is a relatively mild disease in adults but has a significant mortality rate in infants. Until immunization was introduced in the 1930s, whooping cough was one of the most frequent and severe diseases of infants in the United States.
Pathogenesis
The disease pertussis has two stages. The first stage, colonization, is an upper respiratory disease with fever, malaise and coughing, which increases in intensity over about a 10-day period. During this stage the organism can be recovered in large numbers from pharyngeal cultures, and the severity and duration of the disease can be reduced by antimicrobial treatment. Adherence mechanisms of B. pertussis involve a "filamentous hemagglutinin" (FHA), which is a fimbrial-like structure on the bacterial surface, and cell-bound pertussis toxin (PTx). Short range effects of soluble toxins play a role as well in invasion during the colonization stage.
The second or toxemic stage of pertussis follows relatively nonspecific symptoms of the colonizaton stage. It begins gradually with prolonged and paroxysmal coughing that often ends in a characteristic inspiratory gasp (whoop). To hear the characteristic sound of whooping cough click whoop.wav (whoop.wav is copyright of Dr Doug Jenkinson, Nottingham, England. www.whoopingcough.net). During the second stage, B. pertussis can rarely be recovered, and antimicrobial agents have no effect on the progress of the disease. As described below, this stage is mediated by a variety of soluble toxins.

{kind=link}
{kind=link}
{kind=link}